Книга белый пароход читать онлайн
Сюжет Основа повести разворачивается вокруг мальчика, живущего среди чужих ему людей, где единственным родным (как по...
В естественных условиях клетки миокарда постоянно находятся в состоянии ритмической активности. В период диастолы мембранный потенциал покоя клеток миокарда стабилен - минус 90 мВ, его величина выше, чем в клетках водителей ритма. В клетках рабочего миокарда (предсердий, желудочков) мембранный потенциал, в интервалах между следующими друг за другом ПД, поддерживается на более или менее постоянном уровне.
Потенциал действия в клетках миокарда возникает под влиянием возбуждения клеток водителей ритма, которое достигает кардиомиоцитов, вызывая деполяризацию их мембран (рисунок 3).
Потенциал действия клеток рабочего миокарда состоит из фазы быстрой деполяризации (0 фаза), начальной быстрой реполяризации (1 фаза), переходящей в фазу медленной реполяризации (фаза плато, или 2 фаза) и фазы быстрой конечной реполяризации (3 фаза) и фазы покоя -- (4фаза).
Фаза быстрой деполяризации создается активацией быстрых потенциалозависимых натриевых каналов, обеспечивающих резкое повышение проницаемости мембраны для ионов натрия, что приводит к возникновению быстрого входящего натриевого тока. Мембранный потенциал уменьшается от минус 90 мВ до плюс 30 мВ, т.е. во время пика происходит изменение знака мембранного потенциала. Амплитуда потенциала действия клеток рабочего миокарда составляет 120 мВ.
При достижении мембранного потенциала плюс 30 мВ инактивируются быстрые натриевые каналы. Деполяризация мембраны вызывает активацию медленных натрий-кальциевых каналов. Поток ионов Са 2+ внутрь клетки по этим каналам приводит к развитию плато ПД (фаза 2). В период плато клетка переходит в состояние абсолютной рефрактерности.
Затем происходит активация калиевых каналов. Выходящий из клетки поток ионов К + обеспечивает быструю реполяризацию мембраны (фаза 3), во время которой медленные натрий-кальциевые каналы закрываются, что ускоряет процесс реполяризации.
Реполяризация мембраны вызывает постепенное закрывание калиевых и реактивацию натриевых каналов. В результате возбудимость миокардиальной клетки восстанавливается -- это период так называемой относительной рефрактерности.
Конечная реполяризация в клетках миокарда обусловлена постепенным уменьшением проницаемости мембраны для кальция и повышением проницаемости для калия. В результате входящий ток кальция уменьшается, а выходящий ток калия возрастает, что обеспечивает быстрое восстановление мембранного потенциала покоя (фаза 4) .
Способность клеток миокарда в течение жизни человека находиться в состоянии непрерывной ритмической активности обеспечивается эффективной работой ионных насосов этих клеток. В период диастолы из клетки выводятся ионы Na + , а в клетку возвращаются ионы К + . Ионы Са 2+ , проникшие в цитоплазму, поглощаются эндоплазматической сетью.
Ухудшение кровоснабжения миокарда (ишемия) ведет к обеднению запасов АТФ и креатинфосфата в миокардиальных клетках, в результате, работа насосов нарушается, вследствие этого, уменьшается электрическая и механическая активность миокардиальных клеток.
Потенциал действия и сокращение миокарда совпадают во времени. Поступление кальция из наружной среды в клетку создает условия для регуляции силы сокращения миокарда.
Удаление кальция из межклеточного пространства приводит к разобщению процессов возбуждения и сокращения миокарда. Потенциалы действия при этом регистрируются почти в неизменном виде, но сокращения миокарда не происходит. Вещества, блокирующие вход кальция во время генерации потенциала действия, вызывают аналогичный эффект. Вещества, угнетающие кальциевый ток, уменьшают длительность фазы плато и потенциала действия и понижают способность миокарда к сокращению.
При повышении содержания кальция в межклеточной среде и при введении веществ, увеличивающих вход ионов кальция в клетку, сила сердечных сокращений увеличивается.
Соотношения между фазами ПД миокарда и величиной его возбудимости показаны на рисунке 5.
Вследствие деполяризации, мембрана кардиомиоцитов становится абсолютно рефрактерна. Период абсолютной рефрактерности у нее продолжается 0,27 с. В этот период мембрана клетки становится невосприимчивой к действию других раздражителей. Наличие длительной рефрактерной фазы препятствует развитию непрерывного укорочения (тетануса) сердечной мышцы, что привело бы к невозможности осуществления сердцем нагнетательной функции.
Фаза рефрактерности несколько короче длительности ПД миокарда желудочков, который длится около 0,3 с.
Длительность ПД предсердий - 0,1 с, столько же длиться систола предсердий.
Период абсолютной рефрактерности сменяется периодом относительной рефрактерности, во время которого сердечная мышца может ответить сокращением лишь на очень сильные раздражения. Он продолжается 0,03 с.
После периода относительной рефрактерности наступает короткий период супернормальной возбудимости, когда сердечная мышца может отвечать сокращением на подпороговые раздражения.
В состоянии покоя внутренняя поверхность мембран кардиомиоцитов заряжена отрицательно. Потенциал покоя определяется в основном трансмембранным градиентом концентрации ионов К+ и у большинства кардиомиоцитов (кроме синусового узла и АВ-узла) составляет от минус 80 до минус 90 мВ. При возбуждении в кардиомиоциты входят катионы, и возникает их временная деполяризация - потенциал действия.
Ионные механизмы потенциала действия в рабочих кардиомиоцитах и в клетках синусового узла и АВ-узла разные, поэтому и форма потенциала действия также различается (рис. 230.1).
У потенциала действия кардиомиоцитов системы Гиса-Пуркинье и рабочего миокарда желудочков выделяют пять фаз (рис. 230.2). Фаза быстрой деполяризации (фаза 0) обусловлена входом ионов Na+ по так называемым быстрым натриевым каналам. Затем, после кратковременной фазы ранней быстрой реполяризации (фаза 1), наступает фаза медленной деполяризации, или плато (фаза 2). Она обусловлена одновременным входом ионов Са2+ по медленным кальциевым каналам и выходом ионов К+. Фаза поздней быстрой реполяризации (фаза 3) обусловлена преобладающим выходом ионов К+. Наконец, фаза 4 - это потенциал покоя.
Брадиаритмии могут быть обусловлены либо снижением частоты возникновения потенциалов действия, либо нарушением их проведения.
Способность некоторых клеток сердца к самопроизвольному образованию потенциалов действия называется автоматизмом. Этой способностью обладают клетки синусового узла, проводящей системы предсердий, АВ-узла и системы Гиса-Пуркинье. Автоматизмобусловлен тем, что после окончания потенциала действия (то есть в фазу 4) вместо потенциала покоя наблюдается так называемая спонтанная (медленная) диастолическая деполяризация. Ее причина - вход ионов Na+ и Са2+. Когда в результате спонтанной диастолической деполяризации мембранный потенциал достигает порога, возникает потенциал действия.
Проводимость, то есть скорость и надежность проведения возбуждения, зависит, в частности, от характеристик самого потенциала действия: чем ниже его крутизна и амплитуда (в фазу 0), тем ниже скорость и надежность проведения.
При многих заболеваниях и под действием ряда лекарственных средств скорость деполяризации в фазу 0 уменьшается. Кроме того, проводимость зависит и от пассивных свойств мембран кардиомиоцитов (внутриклеточного и межклеточного сопротивления). Так, скорость проведения возбуждения в продольном направлении (то есть вдоль волокон миокарда) выше, чем в поперечном (анизотропное проведение).
Во время потенциала действия возбудимость кардиомиоцитов резко снижена - вплоть до полнойневозбудимости. Это свойство называется рефрактерностью. В период абсолютной рефрактерности никакой раздражитель не способен возбудить клетку. В период относительнойрефрактерности возбуждение возникает, но только в ответ на надпороговые раздражители; скорость проведения возбуждения снижена. Период относительной рефрактерности продолжается вплоть до полного восстановления возбудимости. Выделяют также эффективный рефрактерный период, при котором возбуждение может возникнуть, но не проводится за пределы клетки.
В кардиомиоцитах системы Гиса-Пуркинье и желудочков возбудимость восстанавливается одновременно с окончанием потенциала действия. Напротив, в АВ-узле возбудимость восстанавливается со значительной задержкой. Сердце: связь между возбуждением и сокращением.
Клетки сердечной мышцы, как и любой другой возбудимой ткани, поляризованы. Мембрана кардио-миоцитов снаружи заряжена поло-жительно, изнутри – отрицательно. Это обусловлено различным содержанием ионов натрия (Na +) и калия (K +) по обе стороны мембраны – внутри сердечных клеток больше K + , а снаружи - Na + (рисунок 9). В покое мембрана кардиомиоцитов непроницаема для ионов Na + , но частично пропускает ионы K + . В результате процесса диффузии в соответствии с концентрационным градиентом ионы K + выходят из кардиомиоцита, увеличивая положительный заряд на его поверхности У клеток рабочего миокарда потенциал покоя составляет – 90 мВ.
В потенциале действия различают следующие фазы (рисунок 10):
Фаза 0 – деполяризация, которая характеризуется повышением натриевой проницаемости за счет активации быстрых натриевых каналов клеточных мембран. В этот период Na + лавинообразно входит в клетку. Эта фаза заканчивается достиже-нием критического уровня деполяризации, при котором происходит изменение знака мембранного потенциала (с -90 мВ до +30 мВ).
Фаза 1 – быстрая начальная реполяризация – связана с активацией медленных натриевых и кальциевых каналов;
Фаза 2 – медленная реполяризация (плато), характеризующееся дальнейшим повышением входа в клетку ионов кальция (Са 2+). В период плато натриевые каналы инактивируются и клетка находится в состоянии абсолютной невозбудимости или рефрактерности.
Фаза 3 – быстрая конечная реполяризация обусловлена активацией калиевых каналов. В период фазы 3 закрываются кальциевые каналы за счет чего падает кальциевый ток, дополнительно деполяризующий мембрану. Это ускоряет процесс реполяризации;
Фаза 4 – потенциал покоя, в период которого за счет работы калий-натриевого насоса полностью восстанавливается градиент концентраций Na + и K + по обе стороны мембраны. Калий-натриевый насос представляет собой белок встроенный в мембрану, который работает таким образом, что выкачивает из клетки 3 иона Na + и закачивает обратно 2 иона K + .
Билет №2: Физиологическое значение собственных гемодинамических сердечных рефлексов с прессорецепторов устьев полых вен, каротидных синусов и дуги аорты.
Наибольшее значение в регуляции работы сердца имеют собственные рефлексы сердечно-сосудистой системы, которые возникают с прессорецепторов устьев полых вен, дуги аорты и каротидных синусов. Возникающие в этих рецепторах под действием давления крови импульсы поступают в центры кровообращения продолговатого мозга и других отделов ЦНС.
При повышении давления крови в полых венах происходит рефлекторное уменьшение тонуса блуждающего нерва и возрастание тонуса симпатического нерва. Это вызывает ускорение сердечного ритма и увеличение силы сокращений (рефлекс Бэйнбриджа). Благодаря этому рефлексу предсердия и венозная часть системы предохраняются от чрезмерного переполнения при интенсивном притоке крови к сердцу.
Особое значение для регуляции гемодинамики играют рефлексы, возникающие с барорецепторов каротидного синуса и дуги аорты. При повышении системного артериального давления раздражение барорецепторов усиливается, это вызывает рефлекторное увеличение тонуса блуждающих нервов и урежение сердечного ритма. В результате уменьшается сердечный выброс и снижается общее артериальное давление. В случае падения общего артериального давления, например, при кровопотере, раздражение каротидных и аортальных барорецепторов уменьшается, снижается тонус вагуса и его тормозное действие на сердце слабеет. В результате происходит увеличение сердечного ритма и минутного объема крови, что приводит к восстановлению системного артериального давления. Наряду с механорецепторами в рефлекторной регуляции кровообращения принимают участие хеморецепторы указанных зон, которые реагируют на изменения напряжения углекислого газа и кислорода в крови. При снижении напряжения кислорода или повышении углекислого газа происходит рефлекторное учащение работы сердца и увеличение артериального давления.
Билет №3: Артериальное давление: волны 1, 2 и 3 порядка. Систолическое, диастолическое, среднее и пульсовое давление. Величина АД у детей различного возраста. Факторы, способствующие повышению давления по мере взросления организма.
Волны первого порядка (1) - самые мелкие, пульсовые, связаны с сокращением и расслаблением левого желудочка. Волны второго порядка (2), более крупные, объединяющие несколько пульсовых волн, связаны с фазами дыхания. Во время глубокого вдоха, когда внутриплевральное давление становится более отрицательным, внутригрудные сосуды расширяются, в них эффективно подсасывается кровь из предшествующих сосудов и давление в артериальной части снижается. При выдохе, когда внутриплевральное давление возрастает, отток крови из артериальной части уменьшается, и это приводит к временному увеличению давления крови. Волны третьего порядка (3) связаны с естественными колебаниями тонуса сосудодвигательного центра.
Систолическое – 110-125, Диастолическое 60 – 85 мм рт. ст., Пульсовое-40 мм рт.ст., Среднее АД-100 мм рт.ст
Рефлекторные механизмы регуляции уровня артериального давления осуществляются путем изменения работы сердца и величины периферического сопротивления. Основными рефлексогенными зонами, в которых локализованы баро- и хеморецепторы являются разветвления сонной артерии и дуга аорты. У взрослых раздражение прессорецепторов этих зон приводит к снижению артериального давления (депрессорный эффект) за счет усиления тонического влияния блуждающего нерва на сердце и снижения прессорного воздействия сосудосуживающего центра на сосуды.
У новорожденных животных (обезьяны) уже функционируют прессорецепторы синокаротидной зоны. Частота импульсов от них зависит от величины артериального давления, но раздражение нервов, идущих от рецепторов вызывает слабовыраженное снижение ситемного артериального давления. Депрессорный эффект с аортальной рефлексогенной зоны отсутствует. Он появляется позже, к 3-4 месяцам, одновременно с формированием тонической активности блуждающего нерва на сердце.
Нестабильны эффекты с хеморецепторов каротидного тельца на гиперкапнию и гипоксию: они не постоянны, либо очень слабые. Только к концу первого года жизни при раздражении хеморецепторов появляется хорошо выраженное повышение артериального давления. Начинают работать регуляторные механизмы перераспределения кровотока при переходе от покоя к двигательной активности.
Сосудодвигательные реакции на гуморальные раздражители появляются раньше, чем на нервные. Так, еще в периоде внутриутробного развития адреналин суживает прекапиллярные сфинктеры. У новорожденных и детей раннего возраста во много раз выше активность ренин-ангиотензинной системы, чем у взрослых. Полагают, что эта система играет у них немаловажную роль в повышении сосудистого тонуса.
ПодробностиВыделяют два типа потенциала действия (ПД): быстрый (миоциты предсердий и желудочков (0.3-1 м/с), волокна Пуркинье (1-4)) и медленный (SA-пейсмейкер 1 порядка (0.02), AV-пейсмейкер 2 порядка (0.1)).
1) Быстрые натриевые каналы (блокируем тетродотоксином) - клетки миокарда предсердий, рабочего миокарда желудочков, волокон Пуркинье, атриовентрикулярного узла (низкая плотность).
2) Кальциевые каналы L типа (антагонисты верапамил и дилтиазем уменьшают плато, снижают силу серд. сокращения) - клетки миокарда предсердий, рабочего миокарда желудочков, волокон Пуркинье, клетки синатриального и атриовентрикулярного узлов автоматии.
3) Калиевые каналы
а) Аномального выпрямления
(быстрая реполяризация) : клетки миокарда предсердий, рабочего миокарда желудочков, волокон Пуркинье
б) Задержанного выпрямления
(плато) клетки миокарда предсердий, рабочего миокарда желудочков, волокон Пуркинье, клетки синатриального и атриовентрикулярного узлов автоматии
в) формирующие I-тoк
, транзиторный выходящий ток волокон Пуркинье.
4) «Пейсмекерные» каналы, формирующие I f – входящий ток, активируемый гиперполяризацией встречаются в клетках синусного и атриовентрикулярного узла, а также в клетках волокон Пуркинье.
5) Лиганд-зависимые каналы
а) ацетилхолин-чувствительные калиевые каналы встречаются в клетках синатриального и атриовентрикулярного узлов автоматии, клетках миокарда предсердий
б) АТФ-чувствительные калиевые каналы свойственны клеткам рабочего миокарда предсердий и желудочков
в) кальций активируемые неспецефические каналы встречаются в клетках рабочего миокарда желудочков и волокнах Пуркинье.
Особенностью потенциала действия в сердечном мышце имеется выраженная фаза плато, за счет которой потенциал действия имеет такую большую длительность.
1): Фаза «плато» потенциала действия. (особенность процесса возбуждения):
ПД миокарда в желудочках сердца продолжается 300-350 мсек (в скелетной мышце 3-5 мсек) и имеет дополнительную фазу «плато».
ПД начинается с быстрой деполяризации клеточной мембраны (от - 90 мв до +30 мв), т.к. открываются быстрые Na-каналы и натрий поступает в клетку. За счет инверсии мембранного потенциала (+30 мв) быстрые Na-каналы инактивируются и ток натрия прекращается.
К этому времени происходит активация медленных Са-каналов и кальций поступает в клетку. За счет кальциевого тока деполяризация продолжается 300 мсек и (в отличие от скелетной мышцы) формируется фаза «плато». Затем медленные Са-каналы инактивируются. Быстрая реполяризация происходит за счет выхода ионов калия (К+) из клетки по многочисленным калиевым каналам.
2) Длительный рефрактерный период (особенность процесса возбуждения):
Все время пока продолжается фаза «плато» натриевые каналы остаются инактивированными. Инактивация быстрых Na-каналов делает клетку невозбудимой (фаза абсолютной рефрактерности , которая продолжается около 300 мсек).
3) Тетанус в сердечной мышце невозможен (особенность процесса сокращения):
Продолжительность абсолютного рефрактерного периода в миокарде (300 мсек) совпадает с продолжительностью сокращения (систола же6лудочков 300 мсек), поэтому во время систолы миокард невозбудим, не реагирует ни на какие дополнительные стимулы; суммация мышечных сокращений в сердце в виде тетануса невозможна! Миокард – единственная мышца в организме, которая всегда сокращается только в режиме одиночного сокращения (после сокращения всегда следует расслабление! ).
Перевод с английского: канд. мед. наук Горелов В. Г., Добродеев А. С., канд. мед. наук Селезнев M. H., канд. мед. наук Цейтлин A. M., Шатворян Б. P.
Дж. Эдвард Морган-мл., Мэгид С. Михаил
М79 Клиническая анестезиология: книга 2-я.- Пер. с англ. - M.-СПб.: Издательство БРШОМ-Невский Диалект, 2000. 366 с., ил.
В книге рассмотрены физиологические основы проведения анестезии у пациентов с сопутствующими заболеваниями сердечно-сосудистой системы, органов дыхания, нервными и психическими расстройствами, нарушениями водно-электролитного баланса и кислотно-основного состояния. В отдельных главах представлены методы проведения анестезиологического пособия в пред-, интра- и постоперационном периодах при хирургических вмешательствах на сердце и сосудах, легких и трахее, пищеводе, головном и спинном мозге и позвоночнике, почках и других органах мочевыделительной системы. Детально освещены вопросы проведения инфузионной терапии - показания, методы, виды растворов, осложнения и альтернативные варианты.
Для врачей-анестезиологов, реаниматологов, студентов медицинских учебных заведений.
Все права защищены. Никакая часть этой книги не может быть воспроизведена в любой форме или любыми средствами, электронными или механическими, включая фотографирование, магнитную запись или иные средства копирования или сохранения информации, без письменного разрешения издательства.
ISBN 5-7989-0165-3 (Издательство БИНОМ) ISBN 5-7940-0044-9 (Невский Диалект) ISBN 0-8385-1470-7 (англ.)
Издание на русском языке: © Издательство БИНОМ, Невский Диалект, перевод, оформление, 2000.
Original edition copyright
© 1996, All Rights Reserved.
Published by arrangement with the Original Publisher,
Appleton & Lange a Simon & Schuster Company
Раздел IV
Анестезиологическое
Пособие
Физиология кровообращения л
q
и анестезия
Анестезиолог должен иметь фундаментальные знания по физиологии кровообращения, которые необходимы как для понимания научных основ специальности, так и для практической работы. В этой главе обсуждаются вопросы физиологии сердца и большого круга кровообращения, а также патофизиологии сердечной недостаточности. Малый (легочный) круг кровообращения рассматривается в главе 22, физиология крови и обмен веществ - в главе 28.
Система кровообращения состоит из сердца и кровеносных сосудов. Она предназначена для снабжения тканей кислородом и питательными веществами и удаления продуктов метаболизма. Сердце перекачивает кровь через две сосудистые системы. В малом круге кровообращения кровь обогащается кислородом и избавляется от углекислого газа. В большом круге - доставляет кислород к тканям и поглощает продукты метаболизма, которые затем элиминируются через легкие, почки или печень.
Сердце
Анатомически сердце представляет собой единый орган, но функционально оно делится на правый и левый отделы, каждый из которых состоит из предсердия и желудочка. Предсердия служат как проводниками для крови, так и вспомогательными насосами для заполнения желудочков. Желудочки выполняют роль главных насосов, перека-
чивающих кровь. Правый желудочек получает дезоксигенированную кровь из большого круга кровообращения и перекачивает ее в малый круг. Левый желудочек получает оксигенированную кровь из малого круга кровообращения и перекачивает ее в большой круг. Четыре клапана обеспечивают однонаправленный поток крови через каждую камеру. Насосная функция сердца обеспечивается сложной последовательностью электрических и механических явлений.
Сердце состоит из специализированной попе-речнополосатой мышечной ткани, заключенной в соединительнотканный каркас. Клетки сердечной мышцы - кардиомиоциты - подразделяются на предсердные, желудочковые, водителей ритма и проводящей системы. Способность кардиомиоцитов к самовозбуждению и их уникальная организация позволяют сердцу функционировать как высокоэффективному насосу. Последовательные соединения между отдельными кардиомиоцитами (вставочные диски), имея низкое сопротивление, обеспечивают быстрое и упорядоченное распространение электрического импульса в каждой камере сердца. Волна возбуждения распространяется от одного предсердия к другому и от одного желудочка к другому по проводящим путям. Связь между предсердиями и желудочками осуществляется не непосредственно, а через АВ-узел, поэтому возбуждение передается с задержкой. За счет этого происходит наполнение желудочка при сокращении предсердия.
Потенциалы действия кардиомиоцитов
Мембрана кардиомиоцита проницаема для ионов К 4 , но относительно непроницаема для ионов Na". Мембраносвязанная Ка + /К 4 -зависимая АТФ-аза перекачивает ионы K + внутрь клетки, а ионы Na" из клетки (глава 28). Концентрация К 4 внутри клетки выше, чем во внеклеточном пространстве. Концентрация Na", наоборот, выше во внеклеточном пространстве, чем внутри клетки. Относительная непроницаемость мембраны для кальция поддерживает высокий градиент концентрации кальция между внеклеточным пространством и цитоплазмой. Выход K + из клетки по градиенту концентрации приводит к потере суммарного положительного заряда внутри клетки. Анионы не сопровождают ионы К 4 , поэтому возникает электрический потенциал, причем внутренняя поверхность клеточной мембраны заряжается отрицательно по отношению к наружной. Таким образом, мембранный потенциал покоя формируется в условиях равновесия между двумя противоположными силами: движением K + по градиенту концентрации и электрическим притяжением отрицательно заряженным внутриклеточным пространством положительно заряженных ионов К".
В норме мембранный потенциал покоя кардиомиоцита желудочка варьируется от -80 до -90 мВ. Если мембранный потенциал становится менее отрицательным и достигает пороговой величины, то в кардиомиоците, как и в клетках других возбудимых тканей (нерв, скелетная мышца), возникает потенциал действия, т. е. происходит деполяризация (рис. 19-1 и табл. 19-1). Потенциал действия вызывает преходящее увеличение мембранного потенциала кардиомиоцита до +20 мВ. В отличие
от потенциала действия нейрона (гл. 14), в потенциале действия кардиомиоцита за пиком следует фаза плато, которая длится 0,2-0,3 с. Потенциал действия скелетной мышцы и нерва обусловлен лавинообразным открытием быстрых натриевых каналов мембраны, потенциал действия кардиомиоцита вызывается открытием как быстрых натриевых каналов (фаза начальной быстрой репо-ляризации), так и медленных кальциевых каналов (фаза плато). Кроме того, деполяризация сопровождается преходящим уменьшением проницаемости мембраны для калия. В последующем проницаемость мембраны для калия восстанавливается, натриевые и кальциевые каналы закрываются и мембранный потенциал возвращается к исходному уровню.
После деполяризации клетки рефрактерны (невосприимчивы) к деполяризующим стимулам до наступления 4-й фазы. Эффективный рефрак-терный период равен минимальному интервалу между двумя импульсами, вызвавшими распространение возбуждения. В быстропроводящих кар-диомиоцитах эффективный рефракторный период практически равен продолжительности потенциала действия. В медленнопроводящих кардиомио-цитах, напротив, эффективный рефрактерный период может превышать продолжительность потенциала действия.

Сюжет Основа повести разворачивается вокруг мальчика, живущего среди чужих ему людей, где единственным родным (как по...

«Наталья боярская дочь», произведение Карамзина — это яркий пример нового течения, которое использовали писатели конца...

Конвертер длины и расстояния Конвертер массы Конвертер мер объема сыпучих продуктов и продуктов питания Конвертер площади...

Сатирические произведения В. Маяковский создавал на всех этапах своего творчества. Известно, что в ранние годы он...

Изучайте свою армию в обыкновенном сраженииДля прохождения кампании вам будет нужно раз за разом повторять серии из...

В число базовых тем, которыми была пронизана лирика А.А.Блока, сумевших стать выражением его ощущения времени, вошла тема...

Когда-то я во мне жили просто непробиваемые негативные установки, касающиеся зарабатывания денег, поиска работы...
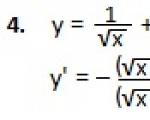
Приведена формула производной суммы и разности функций. Приведено доказательство и подробно разобраны примеры...

Всем известная басня “Кукушка и Петух ” написана Крыловым по конкретному поводу. Ее публикация в сборнике...

Закономерность исторического процесса потребовала усиления феодализации древнекитайского общества. И история...

Значение фразеологизма «ни рыба ни мясо»Рассмотрим довольно распространенный фразеологизм русского языка “ни...

"Пломбированный вагон"Список пассажировСписок взят из газет В. Бурцева «Общее дело» за 14.10.1917 и...

Вы уже знаете, что союз – это служебная часть речи, с помощью которой оформляют связь между частями...

Подданство: Страна: Ошибка Lua в Модуль:Wikidata на строке 170: attempt to index field "wikibase" (a nil...

«Наталья боярская дочь», произведение Карамзина — это яркий пример нового течения, которое использовали...

Конвертер длины и расстояния Конвертер массы Конвертер мер объема сыпучих продуктов и продуктов питания...